
















Glukagoni on peptidihormoni, jota tuottavat haiman alfasolut. Glukagoni lisää glukoosin ja rasvahappojen pitoisuutta verenkierrossa, ja sitä pidetään kehon tärkeimpänä katabolisena hormonina. Glukagonia käytetään myös lääkkeenä useiden terveydentilojen hoitoon. Glukagonin vaikutus on päinvastainen kuin insuliinin, joka alentaa solunulkoista glukoosia. Glukagonia tuotetaan proglukagonista, jota GCG-geeni koodaa.
Haima vapauttaa glukagonia, kun glukoosin määrä verenkierrossa on liian pieni. Glukagoni saa maksan sitoutumaan glykogenolyysiin: muuttamalla varastoitua glykogeenia glukoosiksi, joka vapautuu verenkiertoon. Korkeat verensokeritasot puolestaan stimuloivat insuliinin vapautumista. Insuliini antaa glukoosin imeytyä ja käyttää insuliinista riippuvaisissa kudoksissa. Glukagon ja insuliini ovat siis osa palautesysteemiä, joka pitää verensokeritasot vakaina. Glukagoni lisää energiankulutusta ja kohoaa stressiolosuhteissa. Glukagoni kuuluu hormonien sekretiiniperheeseen.
Glukagonin toiminta
Glukagoni yleensä nostaa veren glukoosipitoisuutta edistämällä glukoneogeneesiä ja glykogenolyysiä. Glukagoni vähentää myös rasvahapposynteesiä rasvakudoksessa ja maksassa. Glukagoni edistää lipolyysiä näissä kudoksissa, mikä saa ne vapauttamaan rasvahappoja liikkeeseen, missä ne voidaan kataboloida energian tuottamiseksi kudoksissa, kuten luurankolihaksissa, tarvittaessa.
Glukoosi varastoidaan maksaan polysakkaridiglykogeenin muodossa, joka on glukaani (glukoosimolekyyleistä koostuva polymeeri). Maksan soluilla (maksasoluilla) on glukagonireseptoreita. Kun glukagoni sitoutuu glukagonireseptoreihin, maksasolut muuttavat glykogeenin yksittäisiksi glukoosimolekyyleiksi ja vapauttavat ne verenkiertoon prosessissa, joka tunnetaan glykogenolyysinä. Kun glukoosivarasto loppuu, glukagon kannustaa maksaa ja munuaisia syntetisoimaan lisää glukoosia glukoneogeneesin avulla. Glukagoni sulkee glykolyysin maksassa aiheuttaen glykolyyttisten välituotteiden siirtymisen glukoneogeneesiin.
Glukagoni säätelee myös glukoosin tuotannon nopeutta lipolyysin avulla. Glukagoni indusoi lipolyysiä ihmisillä insuliinin suppressio-olosuhteissa (kuten tyypin 1 diabetes mellitus).
Glukagonituotanto näyttää olevan riippuvainen keskushermostosta vielä määrittelemättömien reittien kautta. Selkärangattomissa eläimissä silmänvarren poistamisen on raportoitu vaikuttavan glukagonin tuotantoon. Nuorten rapujen silmänvarren leikkaaminen tuottaa glukagonin aiheuttamaa hyperglykemiaa.
Glukagonin vaikutusmekanismi
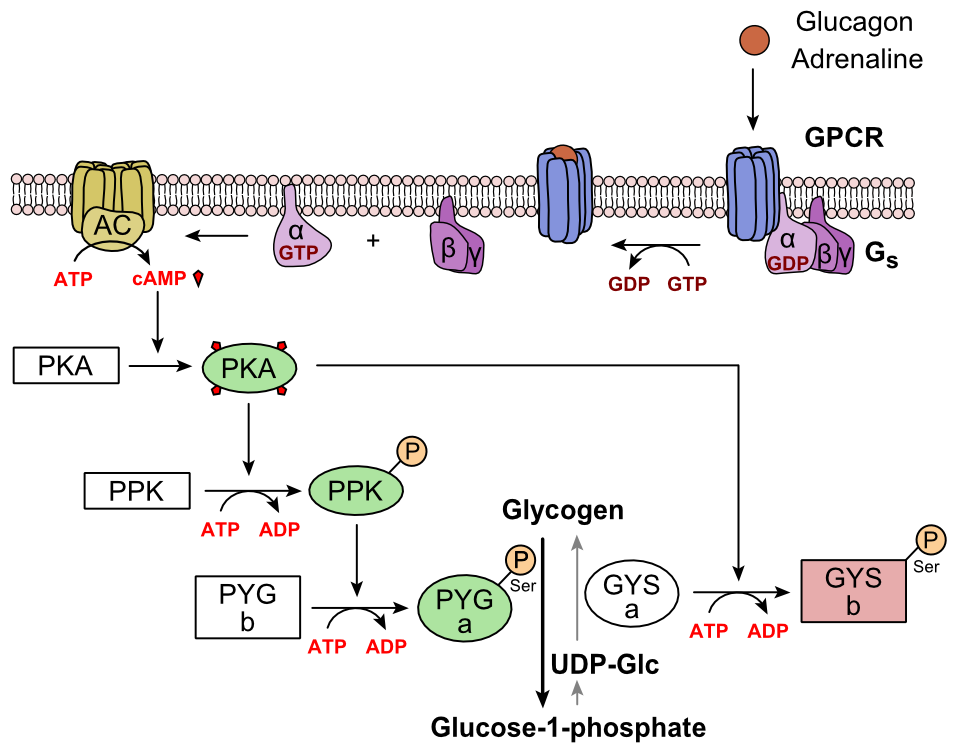
Glukagoni sitoutuu glukagonireseptoriin, G-proteiiniin kytkettyyn reseptoriin, joka sijaitsee solun plasmakalvossa. Konformaation muutos reseptorissa aktivoi G-proteiinit, heterotrimeerisen proteiinin, jolla on a-, p- ja y-alayksiköitä. Kun G-proteiini on vuorovaikutuksessa reseptorin kanssa, se käy läpi konformaationmuutoksen, joka johtaa a-alayksikköön sitoutuneen GDP-molekyylin korvaamiseen GTP-molekyyliin. Tämä substituutio johtaa a-alayksikön vapautumiseen p- ja y-alayksiköistä. Alfa-alayksikkö aktivoi spesifisesti seuraavan kaskadin entsyymin, adenylaattisyklaasin.
Adenylaattisyklaasi valmistaa syklistä adenosiinimonofosfaattia (syklinen AMP tai cAMP), joka aktivoi proteiinikinaasi A: n (cAMP-riippuvainen proteiinikinaasi). Tämä entsyymi puolestaan aktivoi fosforylaasikinaasin, joka fosforyloi sitten glykogeenifosforylaasi b: n (PYG b), muuttamalla sen aktiiviseksi muodoksi, jota kutsutaan fosforylaasi a: ksi (PYG a). Fosforylaasi a on entsyymi, joka on vastuussa glukoosi 1-fosfaatin vapautumisesta glykogeenipolymeereistä.
Esimerkki reitistä olisi, kun glukagon sitoutuu kalvon läpi kulkevaan proteiiniin. Transmembraaniproteiinit ovat vuorovaikutuksessa G12P: n kanssa. Gɑ erottuu Gβ2: sta ja on vuorovaikutuksessa transmembraaniproteiinin adenylyylisyklaasin kanssa. Adenylyylisyklaasi katalysoi ATP: n konversiota cAMP: ksi. cAMP sitoutuu proteiinikinaasi A: han ja kompleksi fosforyloi fosforylaasikinaasia. Fosforyloitu fosforylaasikinaasi fosforyloi fosforylaasia. Fosforyloitu fosforylaasi leikkaa glukoosiyksiköt glykogeenista glukoosi-1-fosfaattina.
Lisäksi maksan glykolyysin ja glukoneogeneesin koordinoitua säätöä säädetään entsyymien fosforylaatiotilalla, jotka katalysoivat voimakkaan glykolyysiaktivaattorin, nimeltään fruktoosi-2,6-bisfosfaatti, muodostumista. Glukagonin käynnistämällä kaskadilla stimuloitu entsyymiproteiinikinaasi A (PKA) fosforyloi myös yhden seriinitähteen bifunktionaalisesta polypeptidiketjusta, joka sisältää sekä entsyymit fruktoosi 2,6-bisfosfataasi että fosfofruktokinaasi-2. Tämä glukagonin käynnistämä kovalenttinen fosforylaatio aktivoi ensimmäisen ja estää jälkimmäisen. Tämä säätelee reaktion katalysoivaa fruktoosi-2,6-bisfosfaattia (voimakas fosfofruktokinaasi-1: n, entsyymin, joka on glykolyysin ensisijainen säätelyvaihe) aktivaattoria hidastamalla sen muodostumisnopeutta, estämällä siten glykolyysireitin virtauksen ja sallimalla glukoneogeneesin hallitsemaan. Tämä prosessi on palautuva ilman glukagonia (ja siten insuliinin läsnä ollessa).
PKA: n glukagonistimulaatio inaktivoi myös glykosyyttisen entsyymin pyruvaattikinaasin maksasoluissa.
Fysiologia
Glukagonituotanto
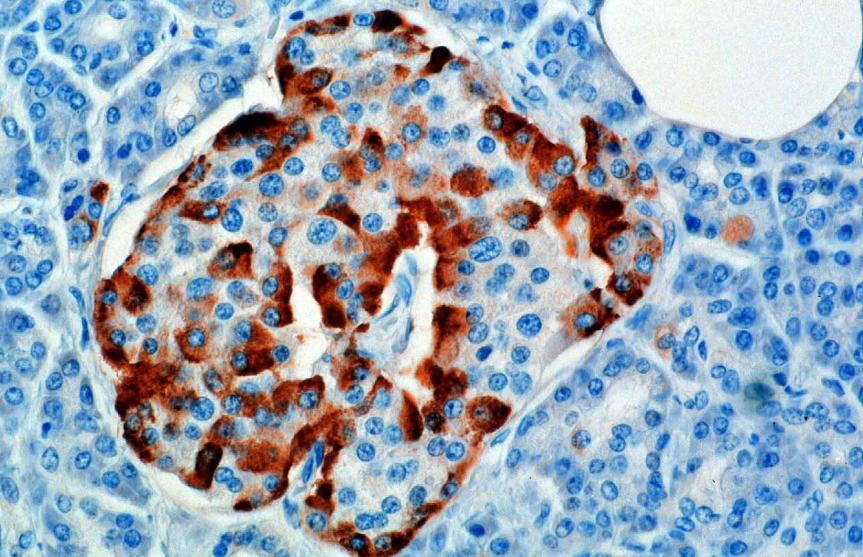
Hormoni syntetisoidaan ja erittyy haiman endokriinisessä osassa sijaitsevien Langerhansin saarekkeiden alfasoluista (a-soluista). Tuotanto, joka on muuten vapaasti käynnissä, tukahdutetaan / säätelee amyliini, peptidihormoni, jota erittyy yhdessä insuliinin kanssa haiman P-soluista. Plasman glukoosipitoisuuksien laskiessa myöhempi amyliinierityksen väheneminen lievittää sen a-solujen tukahduttamista, mikä mahdollistaa glukagonierityksen.
Jyrsijöissä alfasolut sijaitsevat saarekkeen ulkoreunassa. Ihmisen saarekkeen rakenne on paljon vähemmän erillinen, ja alfasolut jakautuvat koko saarekkeelle beetasolujen läheisyyteen. Glukagonia tuottavat myös mahassa olevat alfasolut.
Viimeaikaiset tutkimukset ovat osoittaneet, että glukagonituotanto voi tapahtua myös haiman ulkopuolella, ja suolisto on todennäköisin haiman ulkopuolisen glukagonisynteesin paikka.
Glukagonin säätely
Glukagonin eritystä stimuloivat:
- Hypoglykemia
- Epinefriini (β2-, α2- ja α1-adrenergisten reseptorien kautta)
- Arginiini
- Alaniini (usein lihasta peräisin olevasta pyruvaatin / glutamaatin transaminaatiosta)
- Asetyylikoliini
- Kolekystokiniini
- Mahalaukun toimintaa estävä polypeptidi
Glukagonin eritystä estävät:
- Somatostatiini
- Amylin
- Insuliini (GABA: n kautta)
- PPARy / retinoidi X -reseptorin heterodimeeri.
- Lisääntynyt vapaiden rasvahappojen ja ketohappojen määrä veressä.
- Lisääntynyt urean tuotanto
- Glukagonin kaltainen peptidi-1
Glukagonirakenne
Glukagoni on 29 aminohapon polypeptidi. Sen ensisijainen rakenne ihmisillä on: NH2-His-Ser-Gln-Gly-Thr-Phe-Thr-Ser-Asp-Tyr-Ser-Lys-Tyr-Leu-Asp-Ser-Arg-Arg-Ala-Gln-Asp-Phe-Val-Gln-Trp -Leu-Met-Asn-Thr-COOH.
Polypeptidin molekyylimassa on 3485 daltonia. Glukagoni on peptidihormoni (ei-steroidi).
Glukagoni syntyy proglukagonin pilkkomisesta proproteiinikonvertaasi 2: lla haiman saarekkeen a-soluissa. Suoliston L-soluissa proglukagoni pilkotaan vaihtoehtoisiin tuotteisiin glysentiiniin, GLP-1 (inkretiini), IP-2 ja GLP-2 (edistää suoliston kasvua).
Patologia
Epänormaalisti kohonnut glukagonitaso voi johtua haiman kasvaimista, kuten glukagonoomasta, jonka oireita ovat nekrolyyttinen vaeltava punoitus, alentuneet aminohapot ja hyperglykemia. Se voi esiintyä yksin tai monen tyyppisen endokriinisen neoplasian yhteydessä.
Kohonnut glukagon on tärkein tekijä hyperglykeemisessä ketoasidoosissa diagnosoimattomassa tai huonosti hoidetussa tyypin 1 diabeteksessa. Koska beetasolut lakkaavat toimimasta, insuliinia ja haiman GABA: ta ei enää ole läsnä tukahduttamaan glukagonin vapaana kulkua. Tämän seurauksena glukagonia vapautuu alfa-soluista korkeintaan, mikä aiheuttaa glykogeenin nopean hajoamisen glukoosiksi ja nopean ketogeneesin. Todettiin, että tyypin 1 diabetesta sairastavien aikuisten alaryhmällä kesti keskimäärin 4 kertaa kauemmin lähestyä ketoasidoosia, kun somatostatiinia annettiin (estää glukagonituotantoa) ilman insuliinia. Glukagonin estäminen on ollut suosittu idea diabeteksen hoidossa, mutta jotkut ovat varoittaneet, että näin tekemällä syntyy hauras diabetes potilailla, joilla on riittävän vakaa verensokeri.
Alfa-solujen (ja siten glukagonin) puuttumisen uskotaan olevan yksi tärkeimmistä vaikutuksista verensokerin äärimmäisessä volatiliteetissa täydellisen haimanpoiston yhteydessä.
Historia
1920-luvulla Kimball ja Murlin tutkivat haiman uutteita ja löysivät uuden aineen, jolla oli hyperglykeemisiä ominaisuuksia. He kuvasivat glukagonia vuonna 1923. Glukagonin aminohapposekvenssi kuvattiin 1950-luvun lopulla. Täydellisempi käsitys sen roolista fysiologiassa ja taudeissa saatiin aikaan vasta 1970-luvulla, jolloin kehitettiin erityinen radioimmunomääritys.
.